| 中央水研ニュースNo.31(2003...平成15年3月発行)掲載 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
【研究情報】
海藻の乳酸発酵技術の開発と新産業創出
内田 基晴
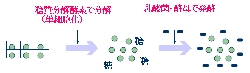
発酵は,古くから食糧生産やエネルギー生産の目的に利用されてきました。海藻の発酵に関しては,これまでエネルギー生産のためのメタン発酵の研究が知られていましたが,食糧生産に役立つ海藻発酵の研究は,ほとんどありませんでした。果たして海藻も野菜や乳のように発酵させることができるのでしょうか?ところで発酵生産物は,植物性素材を発酵させたものと動物性素材を発酵させたものとに大きく分類することができます。例えば身の回りを眺めてみると,植物性素材を発酵させてできたお酒や味噌や納豆,動物性素材を発酵させてできたチーズやヨーグルトなどがすぐ頭に浮かびますが,これらは,ほとんどが陸系の発酵素材です。それでは,海洋系の発酵素材はというと魚醤油やかつおぶしといった動物性のものはありますが,実は海藻のような植物性の発酵素材を造り出す技術は,これまでありませんでした(図1)。本稿では,中央水産研究所において最近開発された海藻の乳酸発酵技術の概要とそれを利用した新産業創出に向けた取り組みを紹介します。
海藻発酵の正体は乳酸発酵 海藻の発酵技術は,偶然の幸運から生まれました。アオサという海藻をセルラーゼで処理して単細胞化した試料を実験に使用した後,その余りを冷蔵庫に1年余りも放置し忘れていたところ,それがワインのようなフルーティーな芳香を有する発酵状態になっていることを偶然観察しました。試しにその一部を同様にセルラーゼで処理した新しいアオサ試料に植え継いだところ,また芳香を有する発酵状態になることが観察され,発酵の元種(スターター)として機能することがわかりました。その後数ヶ月おきに植え継ぎをおこない1年以上にわたり安定に発酵スターターとして機能することを確認したのち,3回および4回植え継いだ海藻発酵試料についてその微生物相を精査したところ,これらの試料中には,1種類の乳酸菌Lactobachillus brevisと2種類の酵母Debaryomyces hansenii var hanseniiおよびCandida zeylanoidesが優占していることがわかりました。このことからここまで官能的な“芳香”だけをたよりに植え継いできた海藻の“発酵”の正体が,どうやらアルコール発酵をともなった乳酸発酵であると推察できました。ここで単細胞化を目的としたセルラーゼによる分解処理は,海藻の糖化即ち発酵基質となる糖の供給に役だっているものと考えられました。これらの点は,後でセルラーゼ添加後に試料中にグルコースの生成と蓄積がおこることおよび発酵の進行によりグルコースの消費と乳酸の生成がおこることで確かめられました。総括すると,海藻発酵試料を植え継ぐ過程で,①セルラーゼ(糖化),②NaCl,③元種の3つの要素を添加することが,発酵を誘起するうえで有効であることがわかりました。分離された乳酸菌であるL. brevisは,陸上で非常によく分離される種類の乳酸菌で,このことからも海藻の発酵は,基本的に野菜の乳酸発酵と同じカテゴリーとみなすことができると考えられました(このことは,実は筆者を,正直がっかりさせたのですけれど)。なお,海藻原料は,生,凍結,乾燥品のいずれでも基質として使用できることがわかりました。 海藻発酵スターターの開発 前述の発酵試料を発酵のための元種(スターター)として使用することは,再現性の観点から問題が残ります。そこで,優占種とされた3種類の微生物の代表分離株,L. brevis B5201株,D. hansenii var hansenii Y5201株,C. zeylanoides Y5206株を元種として使用することを検討しました。その結果,別々に培養した3菌株を混合接種するやり方でも問題なく,アオサの乳酸発酵が再現されることが確認されました。図2に海藻発酵の原理を示しました。さらにこの微生物コンソーシアムをセルラーゼとともに加えると紅藻類,褐藻類,緑藻類,顕花植物などすべての種類の海藻を乳酸発酵もしくはエタノール発酵させることができることがわかりました(表1:前ページ)。ちなみに発酵後に生成する乳酸・エタノールの量から判断すると,顕花植物,緑藻類,紅藻類の順に発酵しやすく,褐藻類が最も発酵しにくい傾向がありました。しかし褐藻類でもワカメは例外的に単細胞化と乳酸発酵が容易であることなどもわかりました。ワカメは規格の揃った乾燥粉末が市販されているため,ワカメ乾燥粉末を基質に使用して海藻葉体を単細胞化しながら乳酸発酵させるために最適なセルラーゼ濃度,NaCl濃度,培養温度等について詳細に調べ,明らかにしました。 微生物相のモニタリング技術の開発 自然の海藻の葉体の表面には,独特の微生物相が形成されていると考えられます。例えばアオサの場合,黄色や橙色の色素を有するFlavobacterium-Cytophaga属の細菌が多く付着していることが知られています。その葉体を放置して腐敗させると悪臭の発生とともにVibrio-Aeromonas属およびPseudoalteromonas属を主体とする微生物相へと遷移することが観察されました。一方,前述のように乳酸発酵をおこさせると悪臭の発生もおこらず,アオサ試料中の微生物相は,乳酸菌と酵母が優占する微生物相となりました。このように微生物相を調べれば,その海藻が発酵状態なのか腐敗状態なのか客観的に評価できます。そこで筆者は,海藻の発酵過程における細菌相の挙動を客観的に把握するため,16S rDNAをターゲットとした菌種特異的プライマーとユニバーサルプライマーを組み合わせてPCR増幅をおこなう方法により,乳酸菌を種レベルまで迅速同定する方法を開発しました。酵母は真核生物であるため18S rDNAをターゲットとして同様に種レベルでの検出法の開発を検討しましたが,18S rDNAの変異が乏しく,種を識別するのに充分な部位がみつからなかったため,より短いランダム合成プライマーを用いるPCR-RAPD法により,迅速同定する方法を開発しました。これにより海藻を発酵させた試料中の微生物相を種レベルで把握することができるようになりました。このモニタリング手法が開発されたことにより,他の乳酸菌と酵母の組み合わせ(例えば食品でよく使われる菌株の組み合わせ)でも海藻発酵に利用できること,乳酸菌の単独接種でもスターターとして機能することを明らかにすることができました。またヤクルト本社との共同研究により,植物系のホモ型乳酸菌を使用することにより,不安定な低塩条件下での安定発酵も実現しました。
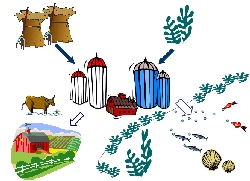
その1 水産飼料素材とマリンサイレージ 海藻発酵素材の利用途として,まず水産飼料としての利用を考え,マリンサイレージという概念を提案しました(図3)。マリンサイレージ(以下MS)とは,“魚介類の栽培を目的として,海藻(藻類)を発酵させて調製した水産飼料”とここでは定義しておきます。筆者らが開発したMSは,海藻の単細胞化と乳酸発酵という二つの技術を核としています。海藻が単細胞化されてあることにより,粒子サイズが直径約10μmとなり微細藻類餌料の代替品としての利用が期待されます。一方,乳酸発酵させていることにより,少なくとも保存性が向上し,常温保存しても長期間腐敗が防げるという利点があります。MSが微細藻類の培養餌料に比べて優れている点は,安定,高密度な培養が低コストでできる点にあります。現場ニーズから考えると未だ充分満足な餌飼料素材が開発されていない二枚貝用飼料としての利用を検討することが特に有望と考えました。まずワカメから調製したMSが,アコヤガイ初期稚貝に対して飼料効果を有することを実証しました。この場合,キートセロスとの併用給餌により飼料効果が相乗的に向上することを観察した点は特に注目されました。一方,MSを単に微細藻類餌料の代替品としてとらえるのではなく,環境問題とリンクさせて利用することを考えることもおもしろいと考えています。アオサは内湾域で大量繁殖する緑藻類で,その藻体の処置に困るケースも多いのですが,栄養塩を吸収している点に着目すれば環境浄化に一役かっているという見方もできます。アオサをMS化し,そのまま浜に戻して二枚貝や動物プランクトンの飼料として機能させ,有用水産物の生産力に結びつけることができれば,環境浄化と食糧生産が同時に達成する循環型システムができあがります。この場合,最初に最終産物(例えばマダイとかヒラメとか)を定めて栽培行為をおこなうのではなく,利用したい一次生産物(この場合アオサ)を決め,いかに効率よく食物連鎖にのせ,またより経済的価値の高い水産物に変換していくかを考えていく点が,従来の増養殖の考えと大きく異なります。
その2 健康食品素材としての利用 海藻を食する習慣は,日本において古くから根付いており,海藻食に対して日本人は漠然とした健康イメージをもっています。一方,乳酸発酵食品というジャンルも,これを食することが健康に良いとする認識が,科学的知見の集積とともにますます高まりつつあります。従って,この二つの要素を併せ持つ海藻発酵素材は,健康機能性を期待させる魅力的な素材といえます。そこでワカメから調製したMSについて,健康機能性を検索するため,ラットを用いた試験により脂質代謝改善作用を有するかどうかを調べました。その結果,MSを1割配合した飼料で3週間飼育したラットの血中及び肝臓中の中性脂質濃度が対照区に比べ,有意に低下することを認めました。また肝臓中のコレステロール値も,対照区に比べ有意に低下していました。発酵処理しないワカメを配合したワカメ区においても同様の傾向が認められましたが,肝臓中の中性脂質濃度は,発酵ワカメ区が,ワカメ区に比べ低い傾向にありました。肝臓酵素の測定から,グルコース-6-リン酸脱水素酵素の活性が有意に低下するなど脂肪酸合成系の代謝が抑制される一方,脂肪酸-CoAのミトコンドリア膜の通過のキャリアであるカルニチン量が有意に増加するなど脂肪酸酸化系の亢進が示唆されました。機能性成分については,現時点では不明ですが,一般成分的には,発酵処理したワカメは,炭水化物成分が減少し,粗タンパク含量が33.1%から39.5%へと増加している点が特徴でした。海藻発酵素材が,健康機能性が期待できる新しい素材であることが示されました。 その3 漬物床としての利用 海藻発酵素材を床としてナスやキュウリを漬け込むとぬか床のように漬け物ができることを観察しています。海藻は,陸上植物に比べ,保水力が高い多糖を多く含むことが特徴です。ヌカ床の場合,一般に理想的な水分含量は55%前後とされ,床の水分含量の管理には気を使わなければなりません。一方,ワカメで調製した漬け物床は,水分含量90%前後であるにもかかわらず,離水がほとんど見られず,水分含量に対する許容範囲が広いと考えられました。また,海藻漬け物床は,高水分であるが故に,高い流動性を有するため,手で掻き回さなくても容器を振るだけで床を攪拌することができます。日本の家庭から漬物床が消えつつある昨今,床の管理に手を汚さないですむ海藻漬物床は,新しい漬け物市場の開拓を予感させます。 新産業創出のためのシナリオ 経済成長の終焉と成熟,漁業資源の枯渇,増養殖事業の成熟と停滞,研究者年齢構成の更新期にあたったこと等が原因してか,水産研究所ではここ20年余り,既存水産業へのテコ入れが中心課題とされ,新産業創出への取り組みが充分ではなかったように思います。まだ“絵に描いた餅”状態ですが,海藻発酵技術を核とした新産業“海洋系植物性基質発酵産業”創出のための私設プログラムを紹介します。まず①農林水産技術会議のパイオニア特別研究により海藻の発酵技術を開発しました。次に②特許出願(3件)をし,技術のプライオリティーを確保しました。それから③学会発表,プレス発表等により成果の情報発信をしました。ここで④低未利用海藻資源を抱える道県(岩手,千葉,北海道)の水産試験場研究員に,研修というかたちで海藻発酵技術の移植をしました。この頃から,民間企業からも問い合わせを頂くようになり,交通整理が必要な状況となりました。そこで想定される新産業をふたつの軸から分類して整理しました。まず利用したい海藻の種類は何かという観点から仕分けしました。これは主に県水試主導の研究開発に適用しました。例えば,岩手県では,ワカメの加工残渣,北海道では,コンブ養殖場周辺の雑海藻,千葉県では,東京湾のアオサ,熊本県では色落ちノリおよび単年性コンブが対象になりそうでした。一方,海藻発酵素材は,何に利用できそうかという観点から,水産飼料,食品,肥料,化粧品という仕分けをし,分野ごとにパートナーを探しました。こちらは主に民間企業が主導で開発研究がスタートしました。実際は10社以上の企業と折衝あるいは情報提供をし,現在も千客万来ウェルカム状態ですが,現在までのところ,水産飼料,食品,肥料の各分野で3社と⑤共同研究契約を締結して研究開発を進めています。ここでの共同研究の成果として,2件の派生特許の共同出願がありました。また⑥特許の使用権契約も別の企業との間で1件成立しました。このように水産研究所のマンパワー不足を補うため,県と民間企業という両輪の軸で共同研究開発を進めることにより,新技術の実用化に取り組んでいます。 上述の①から⑥までを3年間でおこなったため,原著論文の執筆が一部しか完了していないことが宿題として残っています。新産業創出への取り組みと論文重視の(独)水産総合研究センター業務評価軸との間にはギャップがあって,両立はなかなか難しいという言い訳をするのに本稿を使わせて頂きました(笑)。 最後に,海藻発酵素材の健康機能性実証のための動物試験では村田昌一博士(当所利用化学部応用微生物研究室長)に,水産飼料効果実証のための二枚貝飼育試験では故沼口勝之博士(当所海区水産業研究部主任研究官)にご指導頂きましたことを御礼申し上げます。 【参考文献】
(利用化学部 主任研究官)
Motoharu Uchida | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

 中央水研ニュース No.31目次へ
中央水研ニュース No.31目次へ