(左図)28℃で培養した正常な細胞。細胞内に常時発現しているHSC70やアクチンが観察される。(右図)37℃で1 時間高温処理した細胞にはストレスタンパク質HSP群が誘導された。

図2 ストレスタンパク質HSP70の転写調節機構
水中で生活する魚類は、水温、浸透圧、低酸素や汚染物質によって、発生、成長、成熟等の生理現象が強く影響さ れるため、このような環境に適応するための環境順化・適応能を発達させている 1) 。これらの機能は、回遊や行動を特徴づけており、水産資源の再生産と種苗生産技術、とくに、発生、稚仔魚の初期生 残に密接に結びつく研究テーマとして位置づけられる。
適応の細胞生物学
環境への適応は細胞と個体レベルの両面から行われている。筆者らは培養細胞を用いて実験系を単純化することによっ
て、内分泌や脳神経系等の高次の生体調節系に制御されるのではなく、個々の細胞で起きている環境因子の影響とそのメ
カニズムを遺伝子レベルで解析することが可能であると考え、熱ショック、低温、浸透圧変化に対する魚類細胞の応答を
調べている。環境の変化、つまり培養条件を変えることによって誘発される遺伝子をまず見つけ、それに関連する遺伝子
群を芋づる式に単離し、環境応答に関する機能を探るという戦略である。もし新しい遺伝子が見つかれば、培養細胞なら
特定の遺伝子機能を高めたり、抑制する実験系も容易に組めるし、他の生物種の細胞との比較も可能である。そこで得ら
れたデータに基づいて実験魚に応用すればよい。ゼブラフィッシュやメダカのトランスジェニック技術によれば特定の遺
伝子機能を改変した魚類の作出も可能であり、それはバイオテクノロジーにもつながる。
環境適応能に関わる基本的な特徴は、個体そのものだけでなく、培養細胞でも維持されている2
,3)。筆者らと特別研究員の坂本竜哉、今春、北里大学を卒業した田中真帆は協同し
てニジマス発眼胚から細胞株を樹立し、その性質を調べた2)。ニジマスの生育に適し
た温度は遊離細胞の培養条件にも反映されており、0~24℃の幅広い範囲で増殖が可能である。冷水性のため25℃以上では
生存できないニジマス細胞に対して、プラティーなどの熱帯魚の場合、培養細胞の適温は20~30℃であり、低温には弱く
、20℃以下では死んでしまう。また、浸透圧への応答に関わる遺伝子の特性は坂本竜哉特別研究員が中心となって解明さ
れつつある。浸透圧についてはニジマス細胞の場合、200~800ミリオスモル/kgの範囲で生育可能である。低張な環境では
細胞は吸水して膨張し、高張では脱水されて収縮するが、魚類細胞にはこのような浸透圧変化にも適応できる機能がある
らしい。ヒトの細胞が37℃の等張液でしか生育できないのとは異なり、はるかに適応範囲が広く、また魚種ごとに異なる
魚類の環境応答は、細胞生物学の興味深い研究テーマである。
ストレスタンパク質でストレスを測る
ストレス応答には他の生物と同様にストレスタンパク質〔熱ショックタンパク質(heat shock protein, HSP)とも呼ば
れる〕が関与している4)。ストレスタンパク質の一種、HSP70は細胞内での発現レベ
ルが高く、これまで最もよく研究されてきた。合成途上や変性したタンパク質の折りたたみ・修復や膜輸送を助ける分子
シャペロンとしての機能が推定されている。筆者が国内留学で京都大学永田和宏教授のもとで魚類のHSP70遺伝子を初めて
単離して以来、魚類のHSP遺伝子の解析を進めてきた5,6)。
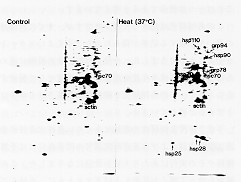
[35S]-メチオニンによって細胞を放射能標識したのち、ストレスタンパク質の誘導を二次元電気泳動法
で観察した。
(左図)28℃で培養した正常な細胞。細胞内に常時発現しているHSC70やアクチンが観察される。(右図)37℃で1
時間高温処理した細胞にはストレスタンパク質HSP群が誘導された。
図2 ストレスタンパク質HSP70の転写調節機構
この研究の端緒は、魚類にとって、急激な温度変化、汚染物質等のストレスの影響をHSP遺伝子の発現レベルで評価 できないだろうか、という点にあった。HSP遺伝子は、プロモーター領域の熱ショックエレメント塩基配列 (nGAAnnTTCn) に熱ショック転写因子heat shock factor (HSF)が結合することによって転写が活性化されるというユニークな特徴を持 つ(図1および2)。魚類の熱ショックプロモーターを上流に連結したガラクトシダーゼあるいはクラゲの蛍光タンパク質遺 伝子を導入したトランスジェニック魚の作出を試みている。表現形質の変換に成功した初のトランスジェニック魚、メラ ニン凝集ホルモン遺伝子の過剰発現によって白色になったメダカを大学時代につくり出した森田貴巳研究員(海洋放射能 研究室)のアドバイスを得て、発生工学の実験系も軌道に乗った。レポータータンパク質の発色によって、さかなのスト レスの度合いを観察することも近い将来可能になるだろう。
ストレスを感じる転写因子
HSFの構造と機能がストレス応答研究の一つの焦点となっている。三量体が転写活性をもつのだが、不活性なHSF前駆体
が細胞内での熱ストレスに応答して、どのように活性化されるのか。HSF自身が細胞内のサーモスタットとして熱ストレ
スを感知し、活性分子へ変化するという仮説の一方で、タンパク質リン酸化のカスケードがHSFの活性化に寄与するとい
う研究もあり、この分野の進展は著しいが、まだ混沌としている。さらに、これまで、ほ乳類ではHSF1とHSF2の二つのタ
イプの存在が知られていたが、最近、ニワトリにはHSF3が、ヒトではHSF4と呼ばれる新しいタイプの転写因子が見つかっ
た。これらのHSFの発現には組織特異性が認められることから、ストレス応答の遺伝子解析はこれまでの培養細胞を用い
た実験系から、個体レベルでの研究にシフトしつつある。変温脊椎動物としての魚類の特徴を活かし、ニジマスとゼブラ
フィッシュを研究材料として、ストレス応答の鍵となるHSFの構造と分子進化、組織分布、ストレスによる活性化様式を
、尾島信彦研究員が中心となって詳細に調べている6)。
4℃で育つニジマス細胞の仕組み
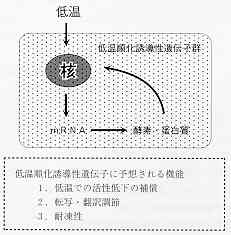
HSP遺伝子の誘導はあらゆる生物に共通して見られるが、魚類で観察される低温への順化応答は、HSPの発現を伴うスト
レス応答とは異なっている。図3に筆者の仮説を図示した 3,7,
8)。低温に伴って誘導される遺伝子群が魚類細胞に存在する。それら遺伝子産物は低温で
比活性の高い酵素・タンパク質アイソフォームであり、細胞の機能低下を補償する。これら遺伝子の転写・翻訳に関与す
るタンパク質因子が低温での遺伝子発現を誘導する。また、0℃前後の氷温では細胞内外の低温障害を防止する作用を示
す物質の存在の可能性も考えられる。
低温順化に関わるタンパク質を探索した。22℃で培養したニジマス細胞を4℃に移し、数日間低温処理すると、細胞 内に特異的に蓄積されるタンパク質が見つかり、酵母のCDC48のホモログであることがわかった3 ,9)。この遺伝子を欠失した酵母は15℃以下では生育できない。このタンパク質は低 温によって低下した細胞分裂能を活性化する役割を果たすことが推定された。さらに、低温処理した細胞で特異的に発現 している遺伝子を探索した結果、フェリチンH鎖遺伝子の転写は低温によって誘導されることが明らかとなった。しかも、 フェリチンH鎖は細胞内に蓄積されることで低温での細胞増殖を促進する効果をもっている7) 。
おわりに
以上のように、筆者らの研究によって、魚類の環境応答のメカニズムに関する新しい事実が明らかにされつつある。魚
類細胞における環境応答には、細胞分裂や細胞骨格、膜機能など細胞機能の変化が関わることが明らかとなってきた。多
様な海洋環境下での生存戦略の解明、ストレス耐性を付与するための育種、魚類細胞を用いる環境影響評価法の開発など
水産分野への応用を目指し、魚類の細胞生物学は進んでいる。
(生物機能部細胞生物研究室)
文献
1) 内田清一郎・菅原浩: "適応の生物学", 講談社 (1977).